組織損傷時や炎症時における末梢での変化
損傷を受けた組織(細胞)からカリウムイオン(K+)・水素イオン(H+)・アデノシン三リン酸(ATP)などが流出します。
また、血小板からはセロトニン、肥満細胞からはヒスタミンが放出され、血漿成分の活性化によってブラジキニン(BK)が作り出されます。
そして、これらの発痛作用(+発痛増強作用)によって痛みが起こります。
BKは発痛物質(+発痛増強物質)であるだけでなく、線維芽細胞・マクロファージ・肥満細胞に作用することでプロスタグランジン(PG)やヒスタミン(His)などの放出を促進させる働きもあります。
PGは単独では発痛作用がないものの、BKの発痛作用を増強する効果(発痛増強作用)があるため、炎症を伴う痛みの感作メカニズムに関与しています(詳しくは末梢神経感作用を参照)。
▼
前述した様々な化学物質により『痛み』だけでなく、局所の血管拡張や血漿成分の滲出により『発赤』や『腫脹』も生じます。
⇒『ブログ:組織損傷時に生まれる発痛物質』も参照
ポリモーダル受容器の刺激により神経炎症が惹起されることで、損傷部周囲の細動脈の拡張と透過性亢進が生じ、損傷部より更に広範囲が赤くなります(=フレア)。
関連記事
⇒『ブログ:ルイスの三重反応』
▼
組織が損傷を受けてから30~60分程度経過すると、好中球やマクロファージ、リンパ球などといった白血球に属する細胞が炎症の主役を演じるようになります。
中でもマクロファージはBKなどに反応し、インターロイキン(IL)や腫瘍壊死因子-αなどの炎症性サイトカインを放出します。そして、この炎症性サイトカインにも発痛増強作用があり、これらもポリモーダル受容器を刺激し、痛みを強めます。
+
ILは線維芽細胞・角質細胞(ケラチノサイト)・シュワン細胞などを活性化させる作用があり、PGや神経成長因子(NGF: nerve growth factor)がこれらの細胞から放出されます。
NGFはポリモーダル受容器の刺激による発痛作用に加えて、神経線維に取り込まれることで後根神経節(DRG: dorsal root ganglion)内の神経細胞体に運ばれ、この細胞体におけるSPやCGRP、TRPV1受容体などの発現増加に影響を及ぼし、結果的に更なる痛みの増強や持続を招いてしまいます。
以下に発物質の発現部位と作用を示します。
| 名称 | 分類 | 発現部位 | 作用 |
|---|---|---|---|
| カリウムイオン(K+) | 電解質 | 損傷細胞 | 発痛作用 |
| 水素イオン(H+) | 電解質 | 損傷細胞 | 発痛作用 発痛増強作用 |
| ブラジキニン(BK) | ペプチド類 | キニノーゲン | 発痛作用 発痛増強作用 |
| サブスタンスP(SP) | ペプチド類 | 1次神経終末(受容器) | 発痛増強作用 |
| CGRP | ペプチド類 | 1次神経終末(受容器) | 発痛増強作用 |
| ヒスタミン(His) | アミノ類 | 肥満細胞 | 発痛作用 |
| セロトニン(5-HT) | アミノ類 | 血小板 | 発痛作用 発痛増強作用 |
| プロスタグランジン(PG) | 脂質 | 細胞膜脂質 | 発痛増強作用 |
| サイトカイン(IL-1など) | その他 | 白血球、マクロファージなど | 発痛増強作用 |
| ATP | その他 | 損傷細胞 | 発痛作用 発痛増強作用 |
一次侵害受容ニューロン
一次侵害受容ニューロンの末梢側末端について
一次侵害受容ニューロン(Aδ線維・C線維)の末梢側末端は自由神経終末と呼ばれる
自由神経終末は『受容器』として機能することで、過剰な関節運動を感知したり、変形に伴う機械適刺激や炎症時の悪物質に反応し、様々な痛みの信号を出しています。
他方で、自由神経終末(のポリモーダル受容器)は『受容器』としてだけでなく、神経ペプチドを放出するといった『効果器』としての機能も備えています。
関連記事
⇒『HP:マイオセラピーの改善機序』
自由神経終末には、高閾値機械受容器・ポリモーダル受容器(+熱・冷侵害受容器)といった『侵害受容器』が、痛覚の受容器として存在する
侵害受容器には、普段はどのような機械的刺激でも反応を示さない『非活動性侵害受容器(silent nociceptor)』といった特殊な受容器も存在します。
この受容器は炎症が生じることによって初めて反応すると言われており、これが痛みの感作に影響していると言われています。
関連記事
⇒『HP:感作(末梢神経感作・中枢神経感作)と脳の可塑的変化』
侵害受容器には、様々な痛み刺激(機械適刺激や化学的刺激)に応答する『受容体』が存在する
神経科学の用語には『受容体』と『受容器』という用語があるが、どちらも『レセプター』という日本語訳なため、混乱しないよう注意が必要です。
痛みに反応する受容器である『侵害受容器』は下記の4つに分類されます。
| 受容器名 | 特徴 |
|---|---|
| 高閾値機械受容器 | 機械適刺激の中でも強い圧迫や切る、刺すなどといった高閾値で、生体にとって侵害域となるもののみに反応し、熱刺激や化学的刺激には反応しない受容器のことをいう。 |
| 熱侵害受容器 | 43°以上の熱刺激に反応する受容器のことをいう |
| 冷侵害受容器 | 15°以下の冷刺激に反応する受容器のことをいう |
| ポリモーダル受容器 | 低閾値から高閾値の機械適刺激、熱指摘、さまざまな侵害性の化学的刺激全てに反応する受容器のことをいう。 ポリモーダル受容器の特筆すべき特徴は、侵害的な刺激を同じ強度で同じ部位(受容野)に繰り返すと、①閾値の低下②刺激に対する反応性の増大③受容野の拡大④自発放電の増大などの現象を示し、これらのことを感作と言う。例えば軟部組織の疼痛が存在した状態でスポーツを行うと、痛みの程度が悪化するばかりでなく、ポリモーダル受容器の感作を引き起こし、それまで痛みを感じていなかった範囲内の動きでも痛みが出現し、筋緊張亢進とともに結果的にパフォーマンスを低下させることになる。 |
上記の侵害受容器の細胞膜には、それぞれの刺激を受け取る以下の一覧表に記載された受容体が存在します。そして、この受容体が活性化すると細胞内情報伝達系が働き、それにより細胞内外のイオン濃度の変化が生じ末梢神経が興奮を起こします。
※以下に記載されている「侵害受容器に関する受容体」と「反応する刺激の種類」の名称は、このサイトのいたる所に登場するので、分からない名称が出てきたら、この表に立ち返ってみてください。
| 受容体 | 反応する刺激の種類 |
|---|---|
| ストレッチ作動性イオンチャネル | 高閾値機械刺激 |
| TRPV1受容体 | 43度以上の熱刺激・カプサイシン・酸(H+) |
| TRPV2~4受容体 | 52度以上の熱刺激 |
| TRPA1受容体 | 17度以上の冷刺激 |
| B2受容体 | ブラジキニン |
| NK受容体 | サブスタンスP |
| IP・EP受容体 | プロスタグランジンI2・E2(PGL2・PGE2) |
| 酸感受性イオンチャネル(ASIC受容体) | 酸(H+) |
| P2X・P2Y受容体 | ATP |
| TrkA受容体 | 神経成長因子(NGF) ※先天性無痛無汗症患者ではTrkA遺伝子に異変があると言われている。 |
~「ペインリハビリテーションの侵害受容器に関する受容体の種類」より~
一次侵害受容ニューロンは、受容体に加わった直接的な刺激(一次的な刺激)のあと、損傷組織から放出される化学的物質による二次的な刺激によって、さらに興奮性が高まります。
(上記の表と重複する記載にはなりますが)侵害受容器の受容体に加わる『一次・二次的な痛み刺激に関与する物質』には下記のような種類が存在します。
- プロスタグランジン(PG:prostaglandin)
- ブラジキニン(BK:bradykinin)
- ヒスタミン(histamine)
- セロトニン(5-HT:5-hydroxy tryptamine)
- アデノシン三リン酸(ATP:adenosine triphosphate)
- 炎症性サイトカイン⇒インターロイキン(IL)・腫瘍性壊死因子(TNFα)
- カプサイシン
- 水素イオン(H+)
- カリウムイオン(K+)
- 神経成長因子(NGF:never growth factor)
- サブスタンスP(SP:substance P)
- カルシトニン遺伝子関連ペプチド(CGEP:calcitonin gene-related peptide)
などなど
一次侵害受容ニューロンについて
一次侵害受容ニューロンにはAδ線維・C線維の2種類が存在し、これらの線維により痛みは脊髄後角まで伝播されます。
- Aδ線維:有髄線維(=伝達早い。一次痛として瞬間的に侵害刺激情報を中枢へ伝える)
深部組織の自由神経終末には高閾値機械受容器・皮膚表面の自由神経終末にはポリモーダル受容器が存在する。 - C線維:無髄線維(=伝達遅い。二次痛として数秒以上かけて侵害重要情報を中枢へ伝える)
自由神経終末にはポリモーダル受容器のみ存在
一次侵害受容ニューロンの終着点(二次侵害受容ニューロンとの中継地点)について
一次侵害受容ニューロンの終着点として、一次侵害受容ニューロンの脊髄側末端は、二次侵害ニューロンの脊髄側末端である脊髄後角のNSニューロン・WDRニューロンとシナプスします。(Aδ線維はNSニューロンのみと、C線維はNSニューロン・WDRニューロンの両方とシナプスします)。
この終着点にて、Aδ線維の脊髄側末端からはグルタミン酸が伝達物質として放出されC線維の脊髄側末端からはグルタミン酸の他に、SP・CGRPが伝達物質として放出されます。
※一次侵害受容ニューロンの脊髄側末端にはカルシウムイオンチャネルが存在します。
そして、活動電位が末端に到達するとチャネルからのカルシウムイオン流入し、末端から伝達物質が放出されます。(つまり、何らかの理由でカルシウムイオンの流入が病的に促進された状態は「シナプス伝導率の長期的な増強」につながるということになります。
※各伝達物質は二次侵害受容ニューロンの下記の受容体へ情報を伝達します。
- グルタミン酸
- ⇒グルタミン酸受容体へ情報を伝達
- SP
- ⇒NK1受容体へ情報を伝達
- CGRP
- ⇒CGB受容体へ情報を伝達
※これ以降の記載は、一次侵害受容ニューロンの脊髄側端末を「シナプス前線維」と記し、シナプス前線維とシナプスする二次侵害受容ニューロンを「シナプス後ニューロン」という用語で統一して記していきます。
一次侵害受容ニューロンの終着点において、シナプス前線維からはグルタミン酸・SP・CGBが放出されます。そしてグルタミン酸は、AMPAと結合することで脱分極し、興奮性シナプス後電位が発生します。
また、AMPA受容体と同じくグルタミン酸の受容体であるNMDA受容体に関しては、通常マグネシウムイオンによって阻害されており、少量のグルタミン酸だけでは活性化されません。
しかし、シナプス前線維からグルタミン酸やサブスタンスPが大量に放出されると、NK1受容体の活性化により、NMDAがリン酸化し、マグネシウムイオンによる阻害がとれてしまいます。
この様にしてNMDA受容体がリン酸化してしまうことでグルタミン酸が結合し、チャネルが開くことでカルシウムイオンがニューロン内に流入してしまいます。
このカルシウムイオンの流入により下記のようなことが生じ、痛覚過敏(中枢性感作)が生じます。
- カルシウムイオンがシナプス後ニューロン内に流入することにより脱分極レベルが増加し、大きな興奮が起こってしまう。
※そして、刺激の反復によって更なる興奮増大につながり「ワインドアップ」という中枢性感作が生じてしまいます。 - カルシウムイオンがシナプス後ニューロン内に流入することにより、PGや一酸化窒素(NO)が産生されます。そして、NOはシナプス後ニューロンの細胞膜から出てシナプス前線維に作用することで、シナプス前線維へのカルシウムイオンの流入を促進してしまう。カルシウムイオンの流入は、シナプス前線維における神経伝達物質の放出の促進につながってしまうため、「シナプス伝導率の長期的な増強」という中枢性感作における機序の一つとされています。
何度も記載してきましたが、この終着点で生じる感作を『中枢性感作』と呼びます。
※「中枢感作」や「中枢神経感作」とも呼ばれます。
もう少し易しく記載された記事、あるいは、興奮性シナプス後電位などの「脊髄後角におけるシナプス後電位」に関しては以下を参照してください。
⇒『ブログ:中枢性感作とは?脊髄後角で起こること』
⇒『ブログ:シナプス後電位』
二次侵害受容ニューロン
一次侵害受容ニューロンによって脊髄後角に入った興奮を、脊髄→延髄→中脳→視床へ伝える神経のことを、『二次侵害受容ニューロン』と呼びます。
NSニューロンとWDRニューロン について
脊髄におけるNSニューロンとWDRニューロンは、一次侵害受容ニューロンとシナプスし、二次侵害受容ニューロンの出発点となります。
- NS(特異的侵害受容)ニューロンの特徴
-
- 侵害刺激にのみ興奮する
- 弱い機械適刺激では興奮しない
- 痛みの発生場所を知らせるニューロンであると考えられている
- WDR(広作動域)ニューロンの特徴
-
- 皮膚だけでなく、筋・内臓などの深部組織からの非侵害刺激から侵害刺激に至るまで幅広い刺激に興奮する。
- 刺激強度に伴って興奮性を増す。
- 痛みを起こす刺激の程度・強度を知らせるニューロンであると考えられている。
- 内側系:
-
- 内側脊髄視床路
- 脊髄網様体視床路
※視床からは大脳の島皮質・前帯状回・扁桃体・海馬へ情報を伝達する
- 外側系:
-
- 外側脊髄視床路
※視床からは大脳皮質の体性感覚野へ情報を伝達する
- 外側脊髄視床路
- 内側系の特徴:
- 身体にとっての痛みの意味ならびに「情動」や「認知」の情報を伝達する(おもに二次痛の経路)
※内側系からの刺激は大脳辺縁系に作用することから、痛みに伴うイライラ感・恐怖・不安などの不快な感情変化を引き起こします。更に間脳の視床下部などにも影響を及ぼし、血圧上昇・頻脈・冷汗・顔面蒼白等の自律神経症状を引き起こします。 - 外側系の特徴:
- 主に痛みの「感覚-識別」に関わる情報を伝達する(主に一次痛の経路)
※WDRニューロンは深部組織の刺激によって皮膚に痛みを感じるという関連痛の発生にも関与すると言われています。さらにWDRニューロンは、繰り返し刺激により感受性を増すワインドアップ(wind-up)と、痛み刺激以外の刺激(例えば触刺激など)によっても興奮し、触覚を伝えるAβ線維からの入力を痛みとして中枢へ伝達するようになります。
脊髄にて痛み刺激は左右に交差して上行する
痛み刺激は、一次侵害受容ニューロンによって脊髄後角にシナプスした後、シナプスした側から反対側へ交叉し、脊髄視床路として脊髄前索・側索を上行し、脳幹レベルで内側と外側に分かれます。
そして、内側系・外側系は下記のように呼ばれます。
内側系と外側系の特徴
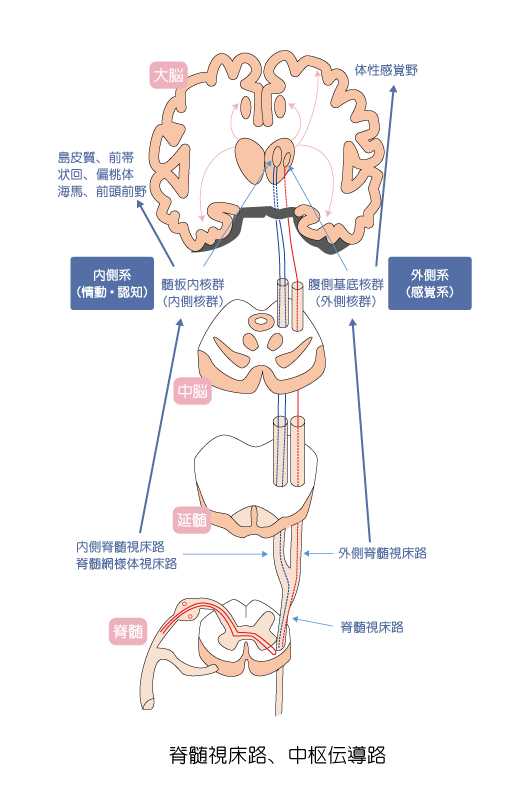
慢性痛は感覚か情動かという議論は以前からありますが、上記の2つの経路が存在し、これらは第三次侵害受容ニューロンにおいても『感覚』、『情動・認知』という異なる場所へいきつくことだけを考えも、両方が作用することは明白です。
セラピストにおいては『痛みは感覚であり、+αの付けたしとして情動も関与している』と考えてしまう側面がありますが、情動が痛みに関与している比重は、意外と大きいかもしれません。
※関連記事
⇒『ブログ:情動と痛み』
⇒『ブログ:感覚・知覚・認知と痛み』
三次侵害受容ニューロン
視床から大脳まで興奮を伝える神経のことを『三次侵害受容ニューロン』と呼びます。
『内側系』の第三次侵害受容ニューロンは視床から大脳の島皮質から前帯状回、前頭前野、扁桃体、海馬に至る系で、身体にとって痛みの意味ならびに「情動」や「認知」の情報を伝達します。
この系は二次痛の経路であり、また大脳辺縁系に作用することから、痛みに伴うイライラ感・恐怖 へ関与するとされています。
また、この経路によって不安感などの不快な感情変化も引き起こし、さらに間脳の視床下部などにも影響を及ぼし、血圧上昇・頻脈・冷汗・顔面蒼白などの自律神経症状を引き起こすとされています。
『外側系』の第三次侵害受容ニューロンは視床から大脳皮質の体性感覚野へ痛みの『感覚-識別』に関わる情報を伝達します。
ただし、体性感覚野が外側系の点というわけではなく、体性感覚野からはさらに補足野、後頭頂葉、島皮質に投射されていきます。
つまり、島皮質は内側系のみならず、外側系の両方から入力を受け、さらに大脳辺縁系を含む広い領域に線維を送っているということになります。
また、体性感覚野から頭頂葉と大脳辺縁系の帯状回、さらに前頭前野を含んだ領域は痛みの認知-評価的側面にかかわっているとされ、帯状回や前頭前野はこれまで経験した痛みの記憶と照合し、痛みを予知した時や注意を集中している時には、痛みの感じ方に影響を及ぼすとされています。
痛み情報の需要に関与する複数の脳領域で、刺激により順番に、または同時に活性化される大脳皮質・大脳辺縁系領域のネットワークの総称は『Painmatrix』と呼ばれています。Pain matrixには、上記の経路で記した「一次体性感覚野」「二次体性感覚野」「島皮質」「前帯状回」「前頭前野」「視床」などが含まれます。
例えば、慢性痛患者と健常者の痛み経験時は、不快感や痛みという感覚経験は同じでも脳内の活動部位が異なることや、視床の活動は減弱または消失する場合があることなど、様々な脳内疼痛処理機構や機能局在とその変化が明らかにされています。
この「視床の活動が減弱または消失している」という点は非常に重要で、これは末梢の痛み情報が視床を経由して脳へ到達していないことを意味し、にもかかわらず痛みが生じているという事実が「脳でつくられた痛み」と表現される所以です。
つまり、同じ痛みでも、慢性痛患者では痛みが持続する中で痛みの中枢神経回路に可塑的変化が生じ、情動や認知にひずみが生じることによって、神経生理学的にも精神心理学的にも修飾され、正常な痛みとは異なる解剖学的局在によって病的な痛みを形成するようになると考えられます。
スポンサーリンク




